History: This four-year-old, spayed female, Chinese crested dog had been
losing weight for one year. No vomiting or diarrhea had been observed by the
owner and the dog had been clinically normal otherwise. There were no apparent
findings in the stomach on gross post mortem examination.
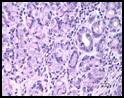
Histopathologic description: There was irregular thickness
of the gastric pyloric mucosa. The lamina propria of the pylorus was markedly
diffusely infiltrated by lymphocytes and plasma cells with scattered lymphoid
follicle formation in the deeper mucosa. The pyloric glandular epithelium was
often attenuated (degeneration) and/or infiltrated by few lymphocytes and
plasma cells. Rarely, pyloric glands were dilated with mucus and scanty cell
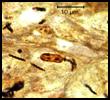
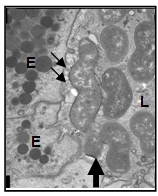
debris. In the pylorus, some gastric pits and glands had varying numbers of
6-10 µm long, spiral, gram-negative bacteria. In the fundus, mucosal
inflammation and bacterial colonization were minimal.
|
Discussion: Severe lymphoplasmacytic gastritis in this dog was
associated with intralesional spiral bacteria. Light and electron microscopic
characteristics of this bacteria are compatible with Helicobacter species. The gastritis might have played a role in the emaciation and weight
loss of this dog.
Genus Helicobacter encompasses gram-negative,
microaerophilic, curved to spiral-shaped bacteria. Chronic gastric
inflammation due to H. pylori has been associated with increased risk of
gastric adenocarcinoma and other malignancies in humans and some animal
species.
Several gastric Helicobacter species have been
isolated from apparently healthy animals or animals with clinical signs of
gastrointestinal problems. In dogs, four gastric Helicobacter species
have been described and the prevalence of these bacteria is generally very high
regardless of concurrent clinical signs. H. (Flexispira) rappini (sheep,
humans, dogs) 4-5 µm, is fusiform, entwined with multiple periplasmic fibers,
and has multiple bipolar sheathed flagella. H. felis (cats, dogs,
humans) is 7-10µm with superficial, sparse periplasmic fibers and multiple
bipolar sheathed flagella. H. heilmannii and H. Gastrospirilium
hominis as synonyms (dogs, cats, humans, nonhuman primates, pigs) is 7-10
µm long, has multiple bipolar sheathed flagella, but lacks periplasmic fibers. H. salomonis (dogs) is 5-7 µm long with tufts of sheathed flagella at
each end. Based on this classification, the majority of bacteria in this dog's
stomach are most consistent with H. felis.
The majority of gastric Helicobacter species
infections in animals has been reported to be asymptomatic; however, some
animals may show intermittent vomiting, weight loss, or diarrhea. Clinical
signs less frequently seen include pica, belching, anorexia, or emaciation.
The mode of transmission of Helicobacter species is yet to be elucidated
although fecal-oral or oral-oral transmission is likely. Until now, there has
been no report pointing to a direct relationship between human infection by
animal Helicobacter species and gastric disorders in humans, even in the
case of H. pylori.
Diagnosis of Helicobacter species infection is
usually made by histologic examination of endoscopic or postmortem stomach
specimens through demonstration of mucosal inflammation accompanied by
organisms. Endoscopic collection of gastric mucus using a brush has been
reported to be equally or more sensitive in detection of the organisms although
the degree of gastric inflammation is difficult to assess. Commercial rapid
urease testing on gastric mucus samples has been used although sensitivity and
specificity are variable. Culture of these fastidious bacteria requires
modification of routine methods and, thus, is not practical. Electron
microscopy and PCR amplification of 16S ribosomal RNA amplicons can be used to
differentiate Helicobacter species.
Pathogenicity of gastric Helicobacter species
relies on their urease production, which increases pH just around the bacterial
wall and provides them with an appropriate milieu. Interestingly, because not
all animals and humans that harbor these bacteria develop clinical signs, both
bacterial virulence and host genetic diversity are thought to be involved in
disease initiation, progression and, in some instances, carcino-genesis. The
reason the pylorus was selectively inflamed and colonized by Helicobacter species in this dog is not clear, although the difference in intragastric distribution
of inflammation and bacterial colonization has often been described in human
and animal infections.
-by Dr. Ikki Mitsui, ADDL Graduate Student
References:
-
Fox JG: 2006. Gastric Helicobacter infections.
In: Infectious Diseases of the Dog and Cat. Ed. Greene CE, 4th ed
W.B. Saunders Company, Philadelphia. PP 343-351.
-
Happonen I et al: 1996. Occurrence and topographical
mapping of gastric Helicobacter-like organisms and their association
with histological changes in apparently healthy dogs and cats. J Vet Med
43:305-315.
-
Jenkins CC, Bassett JR: 1997. Helicobacter infection. The Compendium 19:267-309.
-
Neiger R, Simpson KW: 2000. Helicobacter infection in dogs and cats: facts and fiction. J Vet Intern Med 14: 125-133.
-
Peek Jr RM, Crabtree: 2006. Helicobacter infection and gastric neoplasia. J Pathol 208:233-248.
-
Simpson KW et al: 1999. Helicobacter felis infection in dogs: effect on gastric structure and
function. Vet Pathol 36:237-248.
|